We work on a number of study systems spanning the diversity of acquired metabolism. You can read fun, non-technical summaries of some of our work here! Holly talked about the lab’s work with Quanta Magazine in 2022 and gave a Simons Presidential Lecture in 2025. Our currently funded research projects include:
- Using the marine ciliate genus Mesodinium as a model system for how acquired metabolism can promote energy generation and stabilize microbial communities (supported by the Army Institute for Collaborative Biotechnologies, the Simons Foundation, and the National Science Foundation Division of Molecular and Cellular Biology).
- Understanding the maintenance and importance of fungal species diversity to tree-ectomycorrhizal mutualisms, and measuring the exchange of information between tree seedlings via a network of ectomycorrhizal partners (supported by the Army Institute for Collaborative Biotechnologies and the National Science Foundation Division of Environmental Biology).
- Experimental evolution of marine mixotrophs in response to climate change. If mixotrophs become more heterotrophic under warmer ocean temperatures, they could mediate a positive climate feedback loop (supported by the Hypothesis Fund, the National Science Foundation Bridging Ecology & Evolution program, an NSF CAREER award, and the Simons Foundation).
- Understanding the epigenetic mechanisms that modulate coral bioenergetics. We use mathematical modeling to understand epigenetic-energetic linkages in this model system for acquired metabolism via metabolic mutualism (supported by the National Science Foundation Rules of Life program).
While a few general research themes are described below, one of the great privileges of mathematical biology is that we are often able to work on a wide array of biological questions. Check out our recent research papers to get the best snapshot of our current interests!
Partner diversity in multispecies mutualisms

Towering Douglas-fir trees in Olympic National Park are supported by a huge belowground diversity of ectomycorrhizal fungal partners.
While much mutualism theory has traditionally focused on pairwise interactions, in reality many mutualistic interactions involve guilds comprised of multiple species. How can we explain the persistence of such high levels of diversity? Our group works on two models for multispecies mutualism. First, we study the metabolic mutualism between trees and ectomycorrhizal fungi, which can involve simultaneous association of host trees with dozens of fungal species. Our work in oaks and pines has shown that fungal traits can help trees persist across a gradient of environmental conditions at the landscape scale. We’re now using a combination of field observations, lab experiments, and eco-evolutionary theory to test the hypothesis that temporal fluctuations in environmental conditions select for relaxed tree investment strategies that maintain a wider array of fungal partners. Second, we collaborate with empiricists to develop mathematical models of coral-dinoflagellate symbiosis, the foundation of diverse coral reef ecosystems. We use bioenergetic and evolutionary models to understand how corals manage (or mismanage their partners) and predict the consequences of climate change on this fragile mutualism.
Acquired Photosynthesis in Marine Planktonic Communities
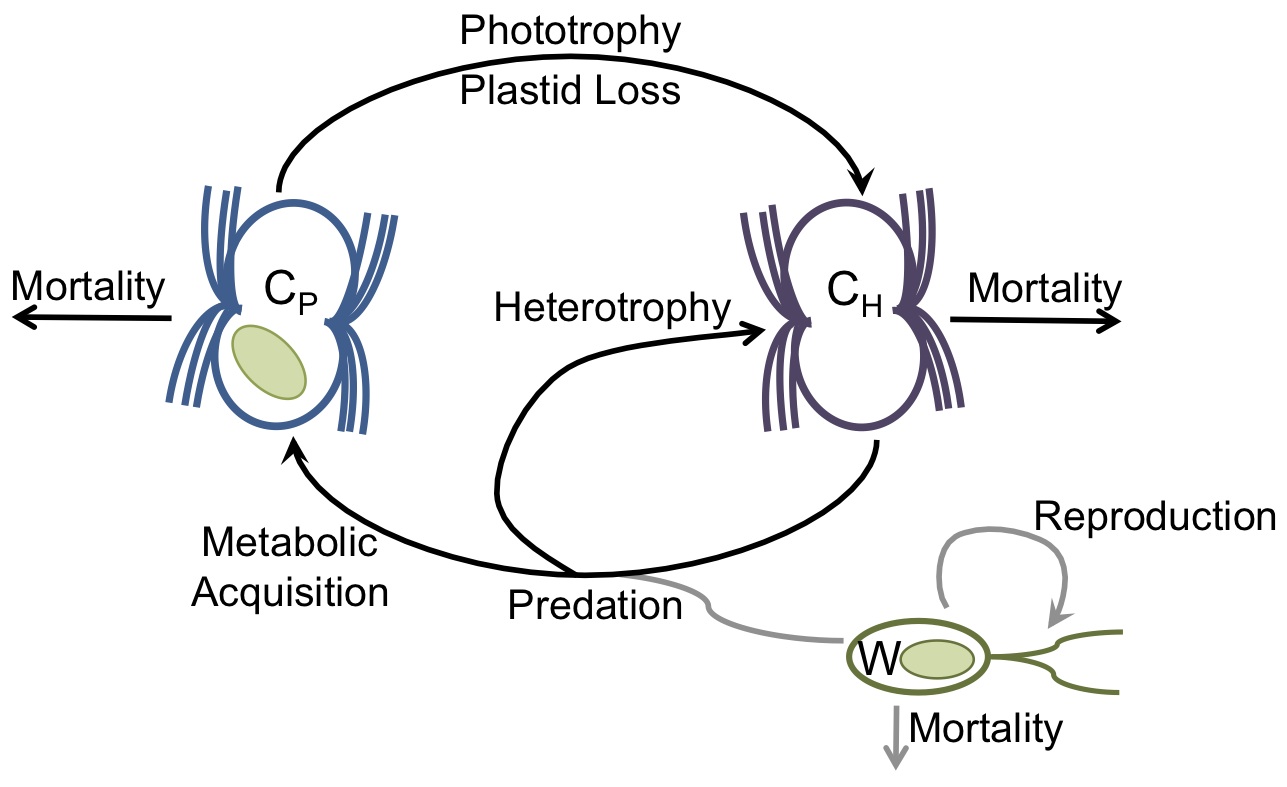
A schematic for acquired photosynthesis, which can be represented with a mathematical model and tested in the laboratory.
Acquired photosynthesis, in which an organism becomes a primary producer by stealing organelles or partnering with an autotrophic endosymbiont, can effectively transform planktonic community members from predators to competitors. In some cases, these acquisitions have become permanent, giving rise to some of the vast diversity of eukaryotic phytoplankton. While a lot of excellent work has been done to identify the genomic signatures of the acquisition of photosynthesis, less is known about the ecological context under which kleptoplastidic (“chloroplast-stealing” lineages evolve), or the selection pressures that cause them to increasingly rely upon photosynthesis and eventually permanently incorporate chloroplasts into their metabolic repertoire. We focus on the genus Mesodinium, a group of ciliates whose members range from totally heterotrophic to almost exclusively photosynthetic, using comparative physiological, ‘omics, and modeling approaches to understand this evolutionary gradient.
Evolutionary response of mixotrophs to changing environmental conditions

Culturing oligotrophic gyre mixotrophs. Image (c) Ken Kostel / WHOI
Mixotrophs (organisms that combine photosynthesis and heterotrophy) play a critical role in the surface ocean by coupling primary production and nutrient remineralization within a single cell. Thus, they can act as carbon sources (via heterotrophy) or sinks (via photosynthesis). But as Earth’s surface waters warm, mixotrophs are expected to become more heterotrophic, which could exacerbate carbon dioxide accumulation in the atmosphere. Our group wants to predict whether this response will be true when mixotrophs are given the opportunity to evolve and adapt to their new environments. We’ve conducted a series of evolution experiments testing mixotroph adaptation, and linking adaptive potential to underlying metabolic tradeoffs. We’re also generally interested in the types of environments that select for mixotrophy, especially environmental heterogeneity (e.g., the frequency of light-dark switching, and the periodicity of nutrient supply).
Optimal control in biological systems

Which partner should I choose? Optimal control theory can help us understand how individuals choose among investments in mutualistic partners, in acquired metabolism, etc.
As a theoretical ecologist, I have particular interests in novel applications of optimal control theory in biological systems. Working with Mike Neubert at WHOI, I use optimal control theory to develop general frameworks for marine reserve design: We’ve shown that profit-maximization can sometimes depend on putting the majority (up to 80%!) of species habitat into reserve. I am also interested in optimal control theory as a way to explore the upper boundaries evolutionary trajectories: As a Ph.D. student, I used an optimal investment framework to study how partner diversity might be maintained in a multispecies mutualism.
<<Top Image: Coral goby surrounded by corals (which acquire photosynthesis through metabolic mutualisms with zooxanthellae), Kapoho Tidepools, Hawaii, August 2008>>
